
Using Optimal Time Varying Inputs to Probe Biochemical Signaling Networks
Joshua F. Apgar, Drew Endy, Forest M. White, & Bruce Tidor
Abstract
Biological signaling networks consist of complex sets of biomolecular interactions that process extracellular signals, such as physical or chemical stimulation, and affect intracellular changes such as protein modifications or changes in gene expression. Complexity in these networks emerges from a variety of factors, including the large number of interacting components, spatial compartmentalization, and feedback loops. Because of this complexity, construction of accurate, mechanistically correct, network models from experimental data remains a significant challenge. One mode of model development involves stimulating a system and measuring the change of network outputs over time. The outputs are often the concentration, activity, or localization of one or more molecular species. The stimulation usually involves step changes of ligand concentration, for example a step up followed by a step down. Candidate models are fit to the data and the best model is selected based on the quality of the fit. Here we explore the extent to which more dynamic stimulation protocols can help improve model selection by revealing hidden complexity in biochemical networks.
A Motivating Example
One example where this idea has been successfully applied is an experiment by Smith-Gill et al. [1]. Smith-Gill was interested in the detailed mechanism of monovalent antibody ligand binding. Preliminary evidence indicated that the binding proceeded in a two-step fashion, whereby the ligand and antibody first came together in a loose encounter complex before proceeding to the fully bound complex. However, this mechanism was difficult to demonstrate because the encounter complex and fully bound complex were indistinguishable by their measurement technique (Surface Plasmon Resonance). Furthermore, the response of the two models to a step up or a step down in ligand was very similar and both could be fit to the data. However, by driving the system with a series of rectangular pulses of varying widths, Smith-Gill was able to find a region where the two mechanisms predicted very different results. For intermediate pulse widths, the two step mechanism showed double-exponential off kinetics corresponding to the off rates of the two species, whereas the one-step model could only produce a single exponential decay. By using this dynamic input signal Smith-Gill was able to more completely explore the dynamics of the system and highlight the existence of an additional species that could not be directly measured.

Preliminary Results
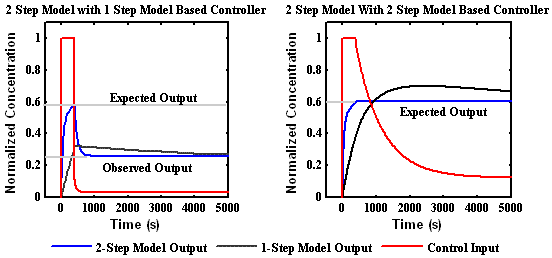
Using the two models proposed by Smith-Gill as a toy system, we have investigated several strategies for designing input signals that can be used to distinguish alternative models. One strategy that has been successful is to construct a signal that will stabilize one model but not the other. The right plot below shows a signal (red) designed to freeze the output of the two step model (blue) but not the one step model (black). Likewise, the left plot shows a signal designed to stop the one step system and not the two step system. By applying these two signals to the system, it is possible to clearly distinguish between the two mechanisms.
References
[1] CA Lipschultz, YL Li, and S. Smith-Gill. Experimental design for analysis of complex kinetics using surface plasmon resonance. In Methods vol. 20, issue 3, pp. 310--318, March 2000.
The Stata Center, Building 32 - 32 Vassar Street - Cambridge, MA 02139 - USA tel:+1-617-253-0073 - publications@csail.mit.edu (Note: On July 1, 2003, the AI Lab and LCS merged to form CSAIL.) |