| Technical Reports | Work Products | Research Abstracts | Historical Collections |
![]()
|
Research
Abstracts - 2006
|
|
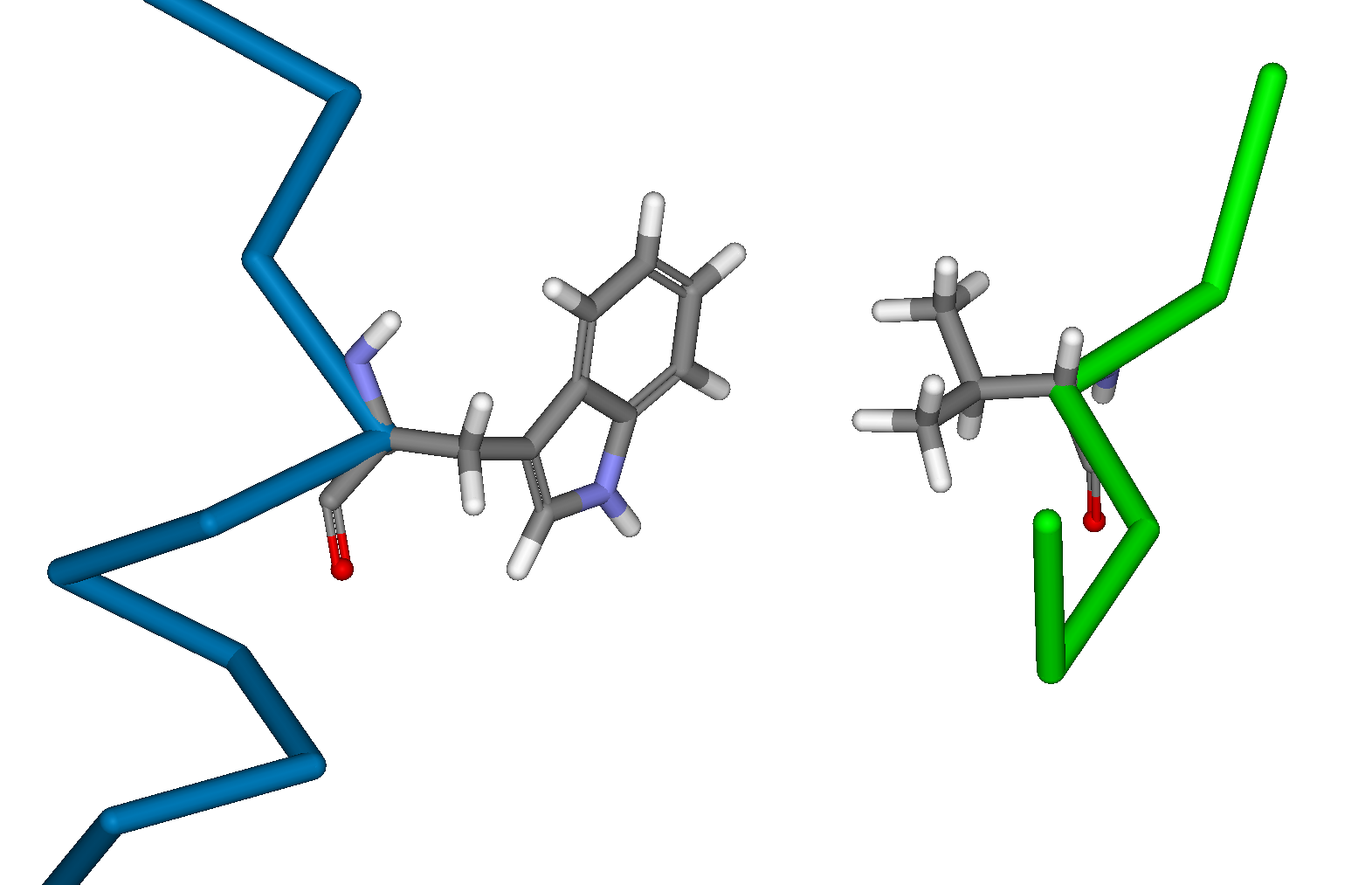
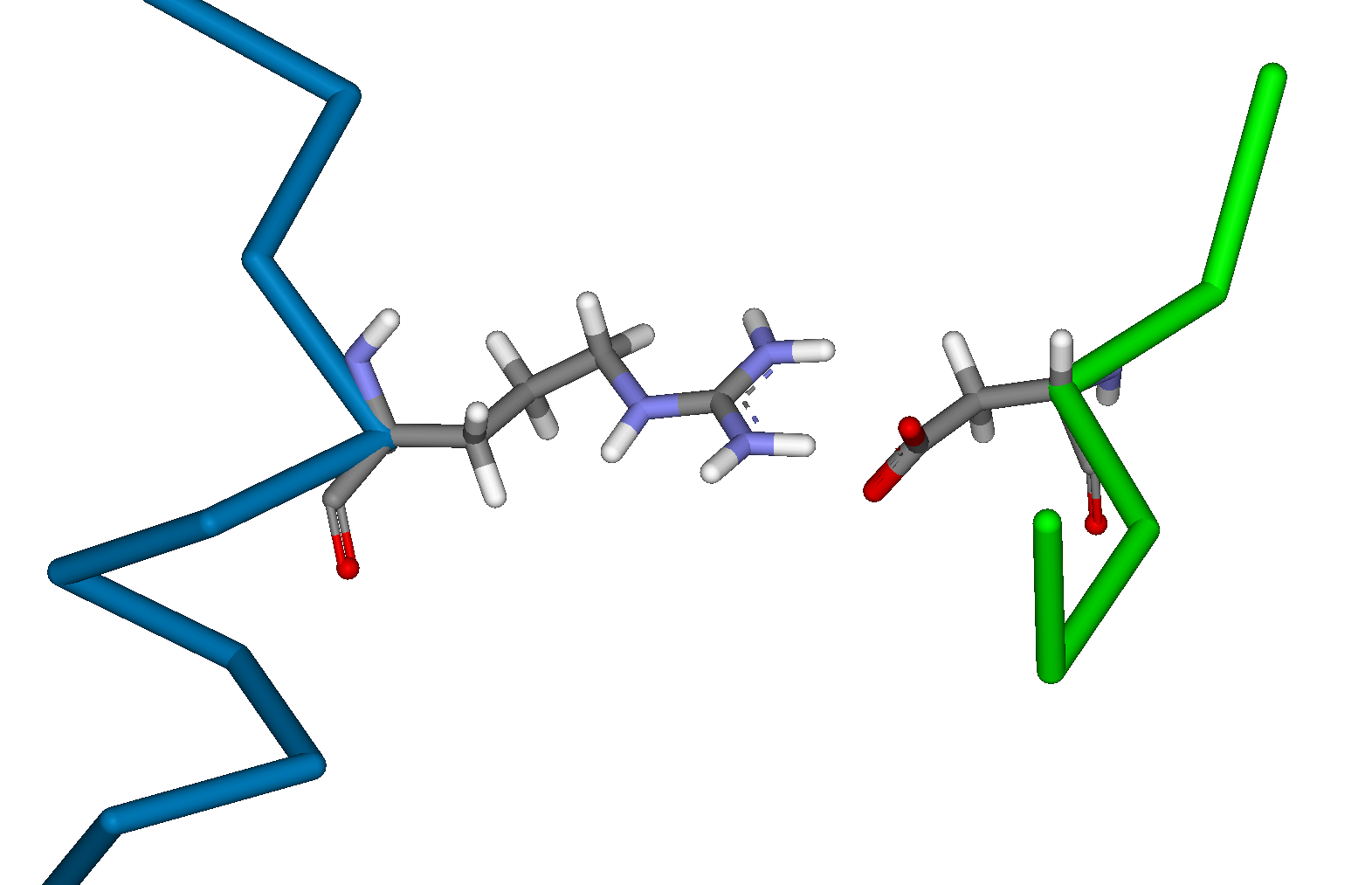
Creation of a Biological Calcium Sensor by the Rational Redesign of CalmodulinDavid J. Huggins, Alan Jasanoff & Bruce TidorIntroductionCalcium is the most abundant mineral in the human body and serves many important biological functions. Within the mammalian brain, the control of calcium-ion flow serves as a basis for neuronal communication and is important in both learning and memory. Calmodulin is a calcium-binding protein that is found in many different cell types and in many different cellular locations. It undergoes a large conformational change upon binding calcium, allowing it to perform its downstream functions. Calmodulin is known to be important in nerve cells, where it is a vital component in long term potentiation. Recent advances in magnetic resonance imagery (MRI) have led to the possibility of using a molecular calcium sensor as a probe for high resolution analysis of neural activity. We have employed computational methods in the task of redesigning Calmodulin and one of its known binding partners (M13) to yield an orthogonal binding pair. This pair could then be used to measure calcium levels in living cells without perturbing the system MethodsThis task involves designing a Calmodulin mutant and an M13 mutant that bind strongly to one another but weakly to the wild type structures. This requires a method for mutating the wild type structures in silico to create realistic structures for the mutants and then an accurate energy function for evaluating these mutants. This allows the comparison of the binding energy of the resulting mutants with the wild type binding energy. This in turn allows the prediction of which mutations are likely to yield the desired result. We initially used the techniques of dead-end elimination [1] in combination with the A* algorithm [2] to perform protein design. These tools have proven to be very useful in reducing the enormous space of protein conformational searches. We built the twenty replacement analogues for each residue in the M13 sequence. The analysis of these replacement analogues directed us to specific sites for mutagenesis. More complex calculations were then performed on these specific sites using CHARMM [3] to optimise the structure of these mutants and calculate van der Waals interactions and differences in solvent accessible surface area. The electrostatic interaction energy and the desolvation penalty were then calculated on the resulting structures using the Delphi program for calculating Poisson-Boltzmann continuum electrostatics [4]. Current ProgressAnalysis of the structure of Calmodulin and the sequences of many of its known binding partners [5] highlighted potentially productive sites for mutation. Calculations were then performed to predict complementary mutations on both sides of the interface that would produce a protein-peptide complex with high binding affinity but minimal cross reactivity with the wild type structures. The most promising mutations were those involving the replacement of key hydrophobic anchor residue of M13 with charged residues and the corresponding mutation of hydrophobic residues of Calmodulin. The wild type and mutant interactions can be seen inFigure 1 for the mutants W4R on M13 and V136D on Calmodulin.
Mutants such as this are now being tested experimentally to investigate their in vitro behaviour. Future PlansThis procedure has the potential to act as a powerful tool in the creation of techniques for the study of this important biological system. We hope to test all our predictions experimentally and design an orthogonal binding pair for this system that can be used experimentally as a biological calcium sensor. This methodology could then be applied to other systems of interest. References:[1] J. Desmet, M. De Maeyer, B. Hazes and I. Lasters. The dead-end elimination theorem and its use in protein side-chain positioning. Nature 356, p539542, 1992. [2] A. R. Leach and A. P. Lemon. Exploring the conformational space of protein side chains using dead-end elimination and the A* algorithm. Proteins 33, p227239, 1998. [3] B. R. Brooks, R. E. Bruccoleri, B. D. Olafson, D. J. States, S. Swaminathan, and M. Karplus. CHARMM: A program for macromolecular energy, minimization, and dynamics calculations. J. Comp. Chem., 4, p187-217, 1983. [4] M. K. Gilson and B. Honig. Calculation of the total electrostatic energy of a macromolecular system: Solvation energies, binding energies, and conformational analysis. Proteins, 4, p7-18, 1988. [5] A. R. Rhoads and F. Friedberg. Sequence motifs for calmodulin recognition. FASEB Journal, 11(5), p331-340, 1997.
|
|||||||||
|